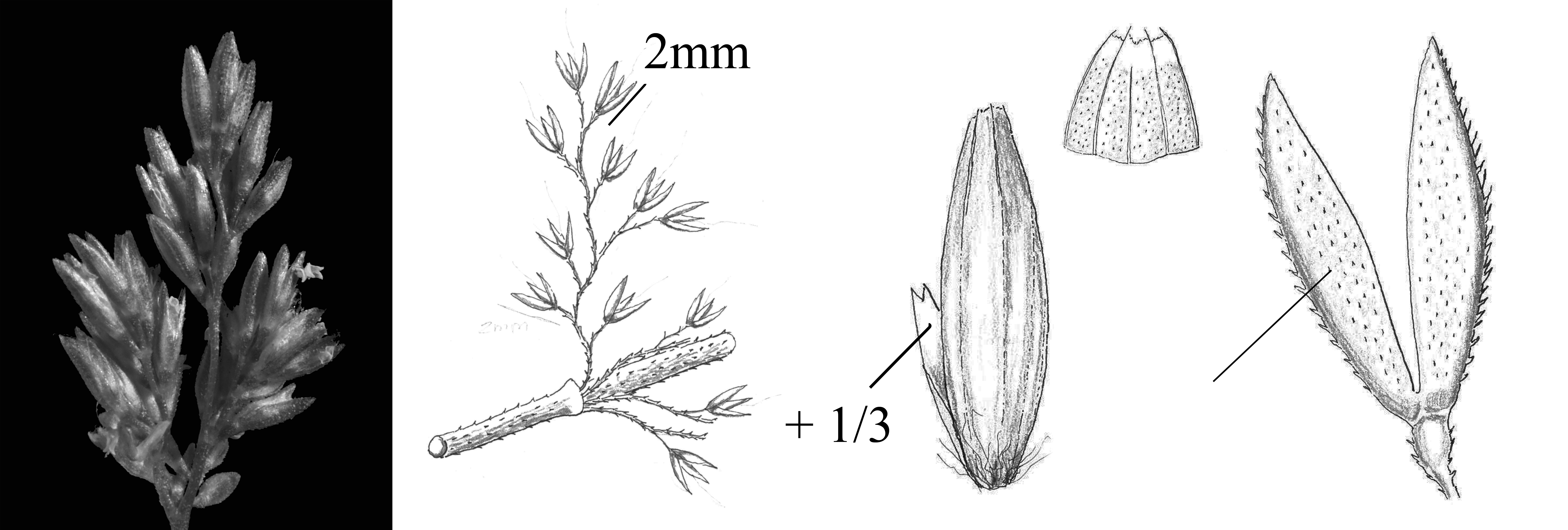
Agrostis se caracteriza por una lema redondeada en el dorso, con arista o sin arista, la pálea diminuta o ausente, la gluma superior con un solo nervio. Las flores son hialinas con un callo con pocos pelos que raramente superan 1/2 de la longitud de la flor.
La tribu Poeae por la filogenia
que se puede obtener del ADN plastidial tiene dos clados bien
marcados: cloroplastos del Grupo 1 (tipo Aveneae) y cloroplastos del
Grupo 2 (tipo Poeae). Hay muchos géneros que se colocan en Aveneae
por su morfología (glumas iguales o que exceden la lema proximal;
espiguillas a menudo de una sola flor; lemas a menudo con aristas
dorsales geniculadas) pero con un tipo de cloroplasto Poeae. De la
misma forma se colocan en Poeae géneros por su morfología (glumas
más cortas o iguales a la lema proximal, espiguillas rara vez con
una sola flor, arista de la lema terminal o subterminal, no
geniculada) que poseen un plástido de tipo Aveneae. Los árboles
filogenéticos que resultan del estudio del ADNnr muestra un grado de
reticulación mayor entre los dos linajes. En este blog seguiremos la
propuesta de Gallher et al. (2022) que divide a la tribu por el tipo
de ADN plastidial.
El género Agrostis pertenece a
subtribu Agrostidinae (Aveneae) que incluye los siguientes géneros.
Calamagrostis (en todo el mundo pero especialmente en Eurasia y
América del Norte); Agrostula (Mediterraneo occidental);
Alpagrostis (Europa occidental); Gastridium ( oeste de
Eurasia) ; Lachnagrostis (África, Asia, Australasia);
Podagrostis (Hemisferio occidental, posiblemente algunas en el
Sudeste Asiático y el este de Asia); Polypogon (en todo el
mundo); Triplachne (Mediterráneo).
Diagrama de relaciones en Agrostidinae modificado de Clayton y Renvoize (1999)
Todas las especies que poseen 14 cromosomas son de origen mediterráneo. Es desde esta región donde se produjo la emigración a otras partes del mundo.
Mapa de distribución de las especies de Agrostis anuales 2n = 14 tomado de A. P. Sokolovskaya "A Caryo-geographical Study of the Genus Agrostis" 1937.
La migración de las formas iniciales (que poseen 14 cromosomas) pudo haber tenido lugar en el periodo posglacial cuando, habiéndose desplazado los elementos florísticos en el período anterior debido al enfriamiento del clima, volvieron tras los glaciares en su retroceso. En su expansión hacia el este de Asia hasta los limites de su área actual , las formas diploides cayeron bajo la influencia de condiciones ambientales muy diferentes a su lugar de origen. Esto provocó la formación de nuevas especies con mayor número de cromosomas. Tal es el tetraploide Agrostis canina 2n = 28 . Una mayor propagación en ambos hemisferios más allá de la zona polar condujo a una diferenciación aún mayor y a la aparición de octoploides 2n = 56 como Agrostis borealis.
Se cree que Agrostis presenta otro centro secundario de especiación en el este de Asia donde se encuentran especies con un elevado número de cromosomas y con unas características morfologicas diferentes a los grupos anteriores, como es el caso de Agrostis capillaris, A. stolonifera 2n =28. (Sokolovskaya, 1937).
Mapa de distribución
del género Agrostis modificado de
http://www.plantsoftheworldonline.org/
Siguiendo clasificación propuesta por Peterson et al. (2020) para el género Agrostis, en el valle del Duero
encontramos las siguientes especies:
A. truncatula, Peterson et al.
(2020) la incluye en el género Agrostula. Este género se diferencia
de Agrostis por que tiene láminas foliares con pilares de
esclerénquima que son continuos entre la superficie adaxial y
abaxial; glumas dorsalmente redondeadas con ápices romos a
truncados; lemas de igual ancho que de largo con ápices ampliamente
truncados, denticulados irregularmente de 5 a 7 dientes, sin arista;
anteras más largas que las lemas y cariopside rugosa-papilosa.
A. truncatula (2n = 14)
Hemicriptófito, en pastos pedregosos acidófilos, silicícola, con
tendencia a ocupar pendientes y zonas erosionadas, de 1300 a 2300 m.
En el Sistema Central, Sierra de la Demanda y S. de Urbión, C.
Cantábrica.
A. curtisii y A. schleicheri, Peterson et
al. (2020) las incluye en el género Alpagrostis. Este género se diferencia
de Agrostis por tener una arista geniculada insertadas en la base y
ápices truncados de la lema con nervios laterales prolongados desde
el ápice en (2) 4 setas; endospermo líquido que se arruga con la
edad.
A. curtisii (2n = 14),
hemicriptófito sin estolones ni rizomas, en bosques acidofilos,
brezales, pastos y grietas de roquedos, silicícola, hasta 1700
m, en la C. Cantábrica y Montes de León.
A. schleicheri (2n = 42),
hemicriptófito sin estolones ni rizomas, en pastos crioturbados y
fisuras kársticas, calcícola, hasta 2400 m, en la C.
Cantábrica.
Las cinco especies que se mencionan a continuación Peterson et al. (2020) las incluye en el género
Agrostis subgenéro Agrostis. Las especies de género Agrostis se
caracterizan por el ápice de la lema entero o dentado con dientes
cortos al final de cada nervio lateral, arista insertada en el tercio
inferior, la mitad o la parte superior del lema pero no basalmente.
Endospermo sólido. El subgénero Agrostis presenta pálea reducida
y malla de tipo I en el lema.
Epidermis del
lema con malla de tipo I en las células largas: (A) A. canina 1100 X
(Widén, 1971) ( (B) A. imberbis 3000X con una escultura similar a
A. Canina (Finot et al. 2013).
A. tileni (2n=14)
hemicriptófito sin estolones ni rizomas ,en pastizales de alta
montaña, silicícola, en la C. Cantábrica y Montes de León.
A. canina (2n = 14),
hemicriptófito con estolones, en ambientes higroturbosos,
silicicola, de 0 a 1000 m, en la C. Cantabrica, S. Demanda y S. de
Urbión, S. Central.

A. hesperica (2n
= 28), hemicriptófito con estolones y rizomas, en prados
higroturbosos, de 0 a 1300 m, en los Montes de León.
A. canina y A.
hesperica se incluyen en el complejo canina que esta formado por un
elevado número de especies distribuido por toda Europa, salvo en la
zona ártica y el sur de las penínsulas mediterráneas, excepto la
Ibérica. En el grupo se ha demostrado la existencia de nº
cromosómico 2n = 14, 28, 42 y 56. En la península Ibérica solo se
han encontrado poblaciones diploides y tetraploides (Romero Gracía,
1985). Las poblaciones tetraploides se extienden por el norte y el
noreste y las diploides preferentemente por el centro, noreste y sur
de la Península. Los diploides presentan solamente estolones y los
tetraploides estolones y rizomas.
A. pourreti (2n
= 14) terófito mediterráneo, en pastizales efimeros sobre suelos
húmedos, en el S. Central y penillanura de Salamanca y Zamora.
A. rupestris (2n
= 14, 28) hemicriptófito sin estolones ni rizomas, en pastos
subalpinos, de 1700 a 2300 m, en la C. Cantábrica, S. Demanda y S.
León, S. Central.
Las cinco
especies siguientes Peterson et al. (2020) las incluye en el género Agrostis
subgénero Vilfa. Las especies de género Agrostis se caracterizan
por el ápice de la lema entero o dentado con dientes cortos al final
de cada nervio lateral, arista insertada en el tercio inferior, la
mitad o la parte superior del lema pero no basalmente. Endospermo
sólido. El subgénero Vilfa presenta palea muy larga de 1/2 a 1/3 de
la longitud del lema (con la excepción de A. tenerrima) y existencia de malla del tipo II-VII.
Epidermis
del lema con malla de tipo VI y VII en las células largas: (A) A.
stolonifera con malla de tipo VI, 1100 X (Widén, 1971) ( (B) A.
stolonifera con malla de tipo VI, 3000X (Finot et al. 2013) (C) malla
de tipo VII, este tipo en realidad representa la ausencia de malla en
el lema, 1100 X (Widén, 1971) (D) malla de tipo VII a 3000X (Finot
et al. 2013).
A. nebulosa (2n = 14),
terófito mediterráneo, en zonas con suelo suelto, arenas con cierto
grado de humedad, de 500 a 900 m, en el Sistema Central.
A. tenerrima (2n = 14).
terófito mediterráneo, en pastos sobre suelo arenoso, hasta 600 m,
en la penillanura de Salamanca.
A. castellana (
2n = 28, 42), hemicriptófito con estolones o rizomas cortos, claros
y orlas de bosques, subatlánticos y mediterraneos, de 400 a 1400m,
en la C. Cantábrica, Montes de León, S. Demanda, S. Urbión y S.
Central.
A. capillaris (2n = 28),
hemicriptófito sin estolones o
rizomas cortos, en prados, matorrales y orlas de bosque, de 0 a 1200
m, en la C Cantábrica, Montes de León y S. Demanda y S.
Urbión.
A. stolonifera (
2n = 28, 42) hemicriptotófito con estolones o rizomas cortos, planta
higrófila que habita en bordes de ríos, balsas, hasta 2200m, en la
C. Cantábrica, Montes de León, S. Demanda, S. Urbión, S. Central y
por el interior de la cuenca.

Agrostis capillaris de Trinius, C.B. "Species graminum" (1828-1836)
Inflorescencia de Agrostis capillaris
Pineda de la Sierra, Sierra de la Demanda (Burgos)







.jpg)