El género Phleum se caracteriza por ser plantas anuales o perennes ; espiguillas unifloras subsésiles, muy comprimidas lateralmente; glumas con tres nervios centrales anastomosados y prolongados en un mucrón o aristadas, en ocasiones con la quilla ciliada que no sobrepasa su longitud; lema 1/2-2/3 de la longitud de las glumas sin aristas; pálea de tamaño similar a la lema.
Estudios moleculares recientes muestran un fuerte apoyo para separar Phleum de Beckmannia y especialmente de Alopecurus (Gillespie et al. 2008). Como resultado, Phleum ahora se trata una vez más en una subtribu distinta, Phleinae y se encuentra cerca de Poa y/o Milium (Boudko, 2014). Alopecurus se diferencia bien de Phleum por el lema aristado, aunque algunos ejemplares de A. arundinaceus son múticos y mantienen en el dorso de la lema una muesca que indicaría el nacimiento de la arista. En la mayoria de las especies ibéricas de Alopecurus falta la pálea mientras que en Phleum es constante.
El género comprende unas 15 especies, siendo el principal centro de distribución geográfica la cuenca mediterránea pero algunos se distribuyen ampliamente en varios hábitats en el norte de Europa, Asia, América del Norte y del Sur (Argentina y Chile). Por ejemplo P. alpinum diploide solo se encuentra en Europa mientras que el tetraploide aparece en Eurasia y Norteamérica. El número base de cromosomas es x = 7.

En el valle del Duero encontramos:
Phleum arenarium (2n = 14) Terófito, en arenales, silicícola, hasta 1800 m. En puntos dispersos en el límite de la provincias de Valladolid y Segovia y en Soria.

Phleum alpinum (2n = 28) Hemicriptófito, en pastos subalpinos, de 1500 a 2600 m. Presente en toda la cordillera Cantábrica y el sierra de la Demanda y picos de Urbión.

Phleum pratense (2n = 2x=14,4x =28, 6x =42, 56) Hemicriptófito, prados, cunetas claros forestales, hasta 1700 m. Por todo el valle del Duero, más abundante que P. phleoides.

Phleum phleoides (2n = 14, 28) Hemicriptófito, en pastos pedregosos soleados, zonas erosionadas hasta 1500 m. Por todo el valle del Duero.

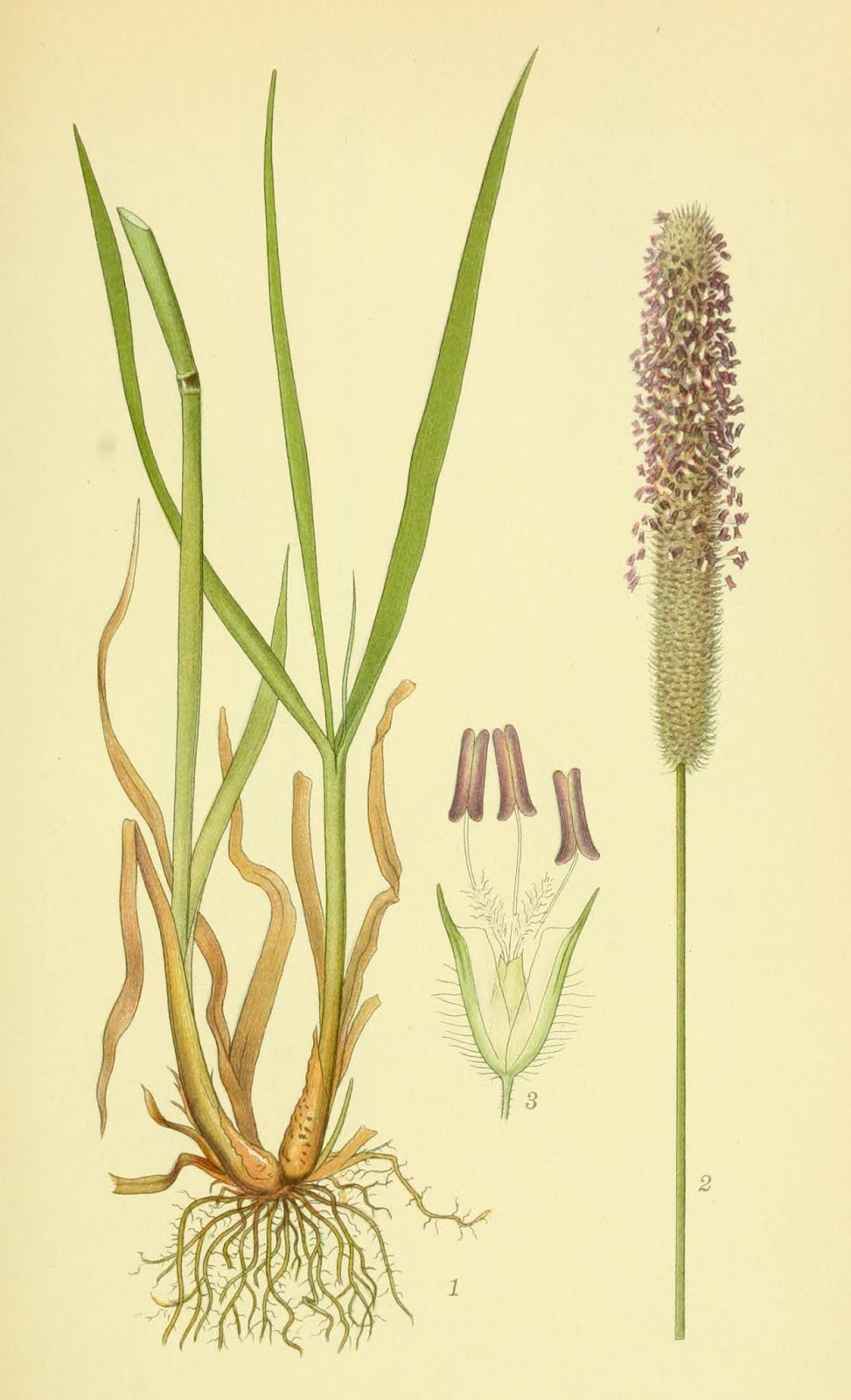
Phleum pratense de Lindman, C.A.M. "Bilder ur Nordens Flora" vol.3 (1922-1926)

Inflorescencia de Phleum pratense



Systematics and karyology of the section Phleum in the genus Phleum
Andrzej Joachimiak & Adam Kula (1997)
En 1997 Joachimiak & Kula escribían acerca de los problemas que crean las divisiones citotaxonómicas en especies que son difíciles de diferenciar por los caracteres morfológicos dentro de la sección Phleum, en el género Phleum. En esta sección, Nordenskiöld (1945) sugirió cuatro especies las cuales diferían en el número de cromosomas: en el grupo P. pratense (P. nodosum, 2n = 2x = 14; P. pratense 2n= 6x = 42) y en el grupo P. alpinum ( P. rhaeticum 2n = 2x = 14 y P. commutatum 2n =4x = 28). En cuanto al aspecto, no se han encontrado diferencias claras que permitan distinguir morfológicamente ejemplares de P.pratense de P. nodosum ( solo la diferencia del mayor tamaño del poliploide). En el grupo P. alpinum, hasta el descubrimiento de un nuevo taxón de Michalski, se pensaba que P. rhaeticum se diferenciaba de P. commutatum sólo en la ausencia de aristas ciliadas en las espiguillas en el último. Pero en 1955 Michalski encontró en los montes Tatra un taxón morfológicamente correspondiente a P commutatum pero con un numero de cromosoma similar a P. rhaeticum.

Espiguilla de Phleum rhaeticum (A) y Phleum commutatum de Andrzej Joachimiak & Adam Kula (1997)
Los estudios realizados por Kula 1996 muestra que existe la posibilidad de resolver el problema mencionado anteriormente estudiando la cantidad de las diferentes fracciones de heterocromatina que presenta el genoma de las especies en cuestión. Kula muestra la presencia de gradientes distintos, posiblemente reflejando la evolución del cariotipo dentro de la sección, lo que arroja luz tanto sobre las relaciones filogenéticas como sobre el origen de los poliploides. Los resultados sugieren un origen alotetraploide de P. commutatum, que probablemente sea un hibrido del taxón de Michalski y de P. rhaeticum, y un origen alohexaploide de P. pratense. No podemos determinar con precisión los progenitores de esta especie. Pero se sostiene a la luz de los estudios realizados que P. pratense contiene 4 genomas de P. nodosum y dos genomas de otra forma. Podrían ser los genomas de P. rhaeticum , o los genomas de un taxón diploide hasta ahora no identificado, estrechamente relacionado con P. nodosum. Esta última posibilidad parece particularmente más interesante, pero el problema requiere más investigación.
En el 2005, Stewart et al., utilizando técnicas de genética molecular fueron capaces de discrimar con precisión las cuatro especies diploides (P. nodosum, P. commutatum (Michalski), P. rhaeticum y finalmente una especie mediterránea poco conocida P. echinatum con un número reducido de cromosomas (2n = 2x = 10). También proporcionaron una idea del origen de algunos poliploides.






.jpg)













