El género Poa se caracteriza por tener plantas esbeltas, la inflorescencia es muy ramificada, con espiguillas al final de delicados pedicelos; tienen espiguillas con más de dos flores (algunas veces una flor en plantas pequeñas); raquilla glabra; glumas ligeramente más cortas que la espiguilla; lemas pubescentes, raramente acuminadas, callo glabro.
El género Poa es el único género dentro de la tribu Poinae. Dentro de esta tribu es problemático ubicar dos géneros: Arctopoa (8 especies, Asia central y oriental, norte de América del Norte) es un hibrido antiguo entre Poa y probablemente algún elemento de Cinninae y Agrostopoa (3 especies, sur de América) posiblemente pertenece a Poa. Todavía los resultados del ADN son provisionales y morfológicamente es más similar a Cinninae (Gallaher et al., 2022).
Se cree que hace 13 millones de años, en el Mioceno, desde el Oeste de Asia (Mediterráneo) se produjo la primera radiación hacia Asia Central y el Este de Asia. Las radiaciones se han ido repitiendo hasta los dos últimos millones de años. Cada radiación se caracteriza por un clado (agrupamiento de varias especies afines filogeneticamente). El género Poa esta formado por cinco clados que corresponde a los cinco subgéneros en los que se divide y que se superponen geográficamente. La primera ramificación corresponde al subgénero Sylvestre que es asiático-americano, representa la radiación más temprana o los restos de dos radiaciones tempranas. Le sigue el subgénero Ochlopoa, la mayoría de las especies que lo forman se encuentran en la región mediterránea europea, unos pocos se extienden hacia el este de Asia, mientras que P. alpina es la única especie autóctona del Nuevo Mundo. Representa la segunda radiación principal que se expandió y se contrajo, una especie sobrevive en el extremo sur de América. Entre esta ramificación y la siguiente se sitúa el subgenero Pseudopoa (sinonimo de Eremopoa) con 7 especies distribuidas desde el mediterráneo oriental hasta el oeste de China. A continuación tenemos el subgenero Stenopoa y Poa centrados en Eurasia-Norteamérica. Estos dos últimos clados alcanzan América del Sur. El subgénero Poa representa la radiación y expansión más reciente del género. Puede que hayan ocurrido múltiples expansiones de especies de este clado durante los periodos glaciales del Pleistoceno (Gillespie et al., 2007).
El género comprende entorno a 500 especies, con número básico n = 7, diploides 2 n = 14 estas son solo el 9% del total de las especies. El resto son poliploides con un contenido bastante alto de hexaploides y decaploides. Además, el cariotipo de una especie P. litorosa consta de 38 subgenomas (2n = 263 a 266). Este es el número cromosómico más alto en el cariotipo de las familia Poaceae conocido hasta ahora (Rodionov et al., 2010) . En las especies de Poa son comunes la hibridación, la apomixia y la introgresión. Los caracteres morfológicos que separan las especies son sutiles y generalmente cuantitativos (Neda Amari, 2016).
El género Poa se cree que tiene un centro de origen Euroasiático a partir del cual se produjo la radiación de las diferentes secciones. Esta idea se tiene en base a las siguientes evidencias: (1) El mayor número de secciones o grupos de Poa se sitúan en Eurasia (2) Siete de las once secciones que incluyen especies diploides están centradas en el oeste de Eurasia. Las otras cuatro secciones con especies diploides son de distribución Ártica y están centradas en Alaska. (3) Entre los países del oeste asiático (Armenia, Libano, Siria...), Irán y Turquía se encuentra las zona que en termino de diversidad de especies de Poa es la más rica (Hoffmann et al. 2013). En 1961, en el Australian journal of Botany, W. Hartley escribía "el género ha alcanzado un alto grado de desarrollo en casi todas las partes del mundo a las que está adaptado climáticamente. La evidencia no permite sacar conclusiones firmes sobre el centro de origen del género, pero tiene un importante centro de diferenciación de especies en la región montañosa y altiplanicie de Asia central".

En el valle del Duero encontramos del subgénero Ochlopoa (P. annua, P. infirma, P. supina, P. bulbosa, P. alpina y P. ligulata); del subgénero Stenopoa (P. minor, P. trivialis, P. compressa y P. nemoralis); del subgénero Poa (P. pratensis, P. cenisia, P. chaixii y P. legionensis) por último sin asignar a ningún subgénero P. flaccidula.
Poa bulbosa (2n = 21, 28, 39, 42) hemicriptófito, en pastos soleados, lugares removidos y nitrogenados, cunetas, hasta 1500 m, por toda la cuenca. La vivíparidad que presenta su inflorescencia es de carácter fisiológico está asociada al pastoreo, especialmente ovino, y es típica de los majadales.

Poa ligulata (2n = 14) hemicriptófito, pastos pedregosos crioturbados, calcícola, de 600 a 1500 m, Cordillera Cantábrica, Sistema Ibérico y Guadarrama.


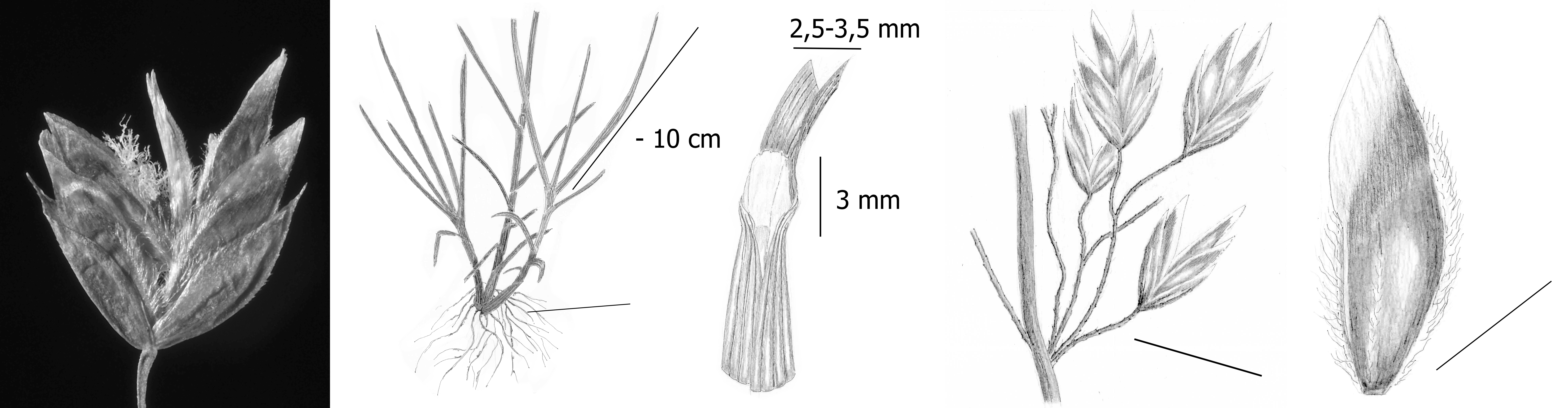

Poa supina (2n = 14) hemicriptófito, pastos encharcados o muy húmedos de montaña, de 900 a 2200 m, puntual por la orla montañosa.



Poa compresa (2n = 42, 45, 49, 56) hemicriptófito, pastos predegosos, graveras, herbazales con cierta humedad, lugares removidos, cunetas, hasta 1600 m, por toda la orla montañosa de la cuenca.

Poa chaixii (2n = 14) hemicriptófito, pastos umbrosos en bosques y prados de montaña, de 1300 a 1950 m, por la Cordillera Cantábrica y norte del Sistema Ibérico.

Poa pratensis (2n = 42, 50-78, 91, 98) hemicriptófito, en dunas y arenales costeros, prados de siega y herbazales, hasta 2000 m, abundante por toda la orla montañosa de la cuenca. Hay distintas razas, generalmente cromosómicas.

TBNC = base del tallo no comprimida
Poa trivialis (2n = 14) hemicriptófito, lugares herbosos con suelo fresco, prados de siega, claros forestales, hasta 1800 m, por toda la cuenca.

Poa nemoralis (2n = 28, 33, 42, 56) hemicritófito, en bosques y sus claros, herbazales sombríos, de 300 a 2200 m, por toda la orla montañosa de la cuenca.

TBNC = Base del tallo no comprimida
Poa cenisia (2n = ?) hemicriptófito, gleras y roquedos de alta montaña, de 1900 a 2200 m, por la Cordillera Cantábrica y Sistema Ibérico septentrional.
Sierra de la Demanda.






Los análisis de los cambios en la vegetación ártica durante el intervalo que va dese hace 50.000 años hasta el presente se han basado principalmente en el polen fósil. Aunque altamente informativos, los registros tienden a estar sesgados hacia productores de polen abundantes como muchas gramíneas (pastos, juncos) y Artemisia, lo que puede oscurecer la abundancia de otras formas como muchas hierbas polinizadas por insectos.
En los últimos años, ha surgido un enfoque complementario que utiliza ADN antiguo de plantas y animales conservado en sedimentos de permafrost. El ADN ambiental puede proporcionar la fracción de la comunidad vegetal no fácilmente identificable por análisis de polen y, en cierta medida, macrofósiles, especialmente en comunidades vegetales dominadas por especies raras o poco representadas en los registros de polen. Para la mayoría de los grupos de plantas, el ADN permite la identificación a niveles taxonómicos más bajos que el polen. Además, los registros de ADN ambiental han demostrado reflejar no solo la diversidad cualitativa, sino también cuantitativa, de plantas y animales sobre el suelo.
Este estudio identifica la vegetación ártica en en tres intervalos distintos:
(1) pre-Último Máximo Glacial (UMG) (50-25 kyr BP), un período de clima fluctuante.
(2) UMG (25-15 kyr BP), un período de condiciones constantemente frías y secas.
(3) post-UMG (15-0 kyr BP), que incluye el interglacial actual, caracterizado por temperaturas relativamente más altas.
Sus resultados muestran que la vegetación pre-LGM estaba dominada por hierbas, mientras que las gramíneas constituían menos del 20% del recuento total de lecturas.
Durante el Último Máximo Glacial (25-15 mil años antes del presente), la diversidad disminuyó notablemente, aunque las hierbas siguieron siendo dominantes. Se mantuvieron proporciones similares de hierbas y gramíneas durante este período. Sus hallazgos contrastan con las reconstrucciones basadas en polen, que han enfatizado la dominancia de las gramíneas en el Ártico no glaciar y regiones adyacentes, particularmente durante el LGM, y están ejemplificadas por el término ampliamente utilizado "estepa mamut".

No hay comentarios:
Publicar un comentario