El género Panicum se caracteriza
por sus inflorescencias en panícula laxa con espiguillas solitarias, de ramas
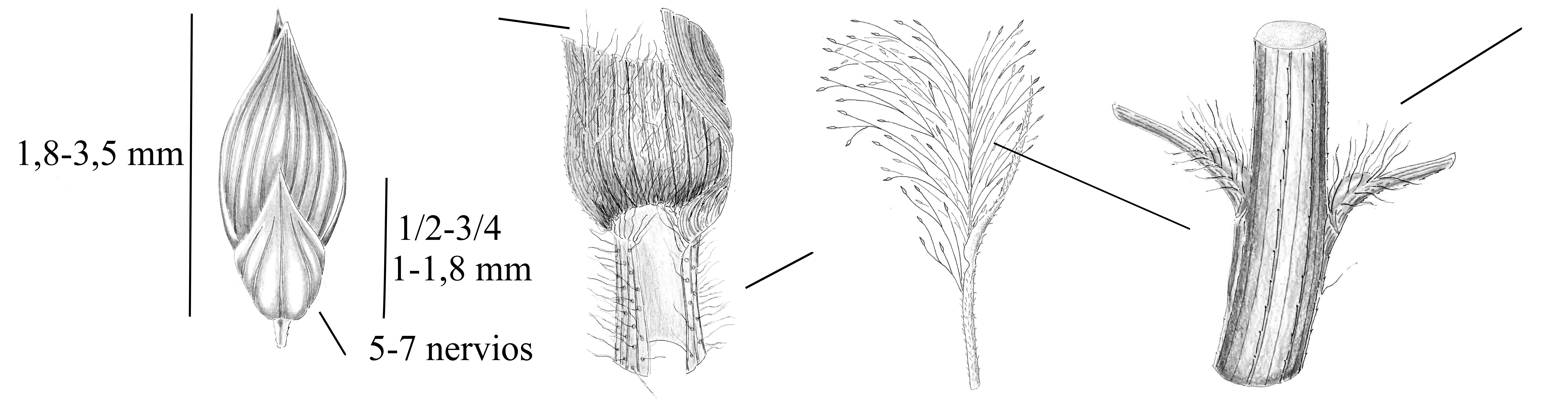
flexuosas; espiguillas comprimidas dorsalmente de 1,8-5 mm, con dos flores, la
inferior masculina o estéril, a menudo representada sólo por el lema, y la
superior hermafrodita; gluma inferior suborbicular mucho más corta que la
superior; lemas muy diferentes en textura, la de la flor superior cartilaginosa y de ápice obtuso, la de la flor inferior blanda y
aguda.

El estudio de Zuloaga et al. (2018) considera que la subtribu Panicinae y Panicum s, str. (71 especies y 7 secciones cuyos nombres aparecen en la parte derecha del gráfico inferior) se diversificaron tempranamente durante el mioceno temprano-medio en la región ancestral Neotropical (Sudamerica), mientras que la subsiguiente diversificación de sus secciones ocurrió principalmente en el Mioceno tardío-Pleistoceno, involucrando múltiples dispersiones a todos los continentes. Esta diversificación tuvo lugar a través de cuatro rutas biogeográficas primarias: 1) Neotrópico-Subsahariano Africa, 2) Neotrópico-América del Norte, 3) África subsahariana-Sudeste de Asia, 4) Australia-Antiguo Mundo. El género Panicum s.l. está formado por unas 600 especies propias de los terrenos intertropicales y subtropicales. El número básico de cromosomas x = 9.


En el valle del Duero
encontramos:
Panicum dichotomiflorum (2n = 4x = 36) Terófito, herbazales nitrófilos, de ordinario con cierta humedad, campos de regadío, arrozales, oriunda de America del Norte, naturalizada en Europa, hasta 800 m. Se presenta de forma puntual en Soria.

Panicum capillare (2n =2x = 18) Terófito, cultivada como ornamental, oriunda de América del Norte, naturalizada en Europa, en cultivos de maíz y terrenos ruderalizados, hasta 700 m. Se presenta de forma puntual en León, Zamora y Salamanca.
Panicum miliaceum (2n = 4x = 36) Terófito, planta cultivada en Europa desde el Neolitico, probablemente procedente de Asia templada o subtropical. La encontramos ocasionalmente asilvestrada en terrenos ruderalizados y cunetas, hasta 900 m. Se distribuye de forma puntual por Segovia, Valladolid y León.






El predominio ecológico de las gramíneas C4 se produjo mucho después de que evolucionaran por primera vez. Según nuestra datación molecular, las primeras gramíneas C4, fueron del grupo Chloridoideae, aparecieron hace 32,0-25,0 millones de años. Sin embargo, la primera evidencia indiscutible de anatomía C4 en el registro fósil (es una gramínea que también pertenece a Chloridoideae) es mucho más reciente y data de hace 12,5 millones de años. La ausencia de fósiles C4 más antiguos sugiere que las gramíneas C4 siguieron siendo raras durante un largo período después de su primera transición desde las C3 o se encontraron en hábitats que no eran propicios para la fosilización. Los isótopos de carbono en paleosuelos indican la presencia de plantas C4 en las grandes llanuras de América del Norte desde el Mioceno temprano (aproximadamente 23 millones de años). Sin embargo, las plantas C4 llegaron a dominar ciertos ecosistemas terrestres solo hace entre 8 y 5 millones de años, lo que pone de relieve la posible importancia ecológica menor, a escala global, de las gramíneas C4 durante los primeros 10 a 20 millones de años de su historia revolucionaria. El surgimiento ecológico de las plantas C4 corresponde a la aparición de biomas dominados por plantas herbáceas, como la sabana. Estos ecosistemas aparecieron y se mantuvieron por perturbaciones externas como el fuego, el pastoreo de herbívoros y los cambios climáticos. En el ambiente recientemente alterado, los árboles C3 fueron reemplazados por especies herbáceas, y las condiciones de luz y temperatura cálidas favorecieron a los taxones C4 sobre los C3. Por tanto, la baja concentración atmosférica de CO2 por sí sola no es suficiente para que las plantas C4 expandan su rango geográfico y ecológico compitiendo con las plantas C3 previamente establecidas. Se necesita una combinación de factores ecológicos para permitir el desarrollo de una flora C4 (por ejemplo, luz intensa y temperatura cálida). Es poco probable que las plantas C4 colonicen hábitats boscosos, donde la luz a nivel del suelo es generalmente extremadamente baja y la temperatura es menor que en hábitats abiertos. Durante su historia temprana las plantas C4 podrían haber permanecido aisladas en algunos hábitats extremos y abiertos donde tenían una ventaja competitiva sobre las plantas C3 hasta que cambios ambientales posteriores les dieron la oportunidad de ascender al dominio ecológico en muchos biomas tropicales y subtropicales (Christin et al. 2008 ).
Existen tres subtipos de la fotosíntesis C4 relacionados con la naturaleza del ácido orgánico transportado a la célula de la vaina del haz y la enzima que cataliza el paso de descarboxilación:
Subtipo PCK donde CO2 se incorpora en las células del mesófilo primero en oxaloacetato y luego como aspartato antes de transportarse a las células de la vaina del haz donde se convierte nuevamente en oxaloacetato por el fosfoenol piruvato citosólico (PEP carboxiquinasa). Comprende géneros característicos de regiones con estaciones secas y húmedas, con altas precipitaciones, en comparación con las especies similares a NAD-ME que son más frecuentes en áreas donde las precipitaciones son inferiores a 30 cm año–1.
Subtipo NADP-ME, el PEP se fija con CO2 para dar oxaloacetato de cuatro carbonos, que se convierte en malato y se transporta a los cloroplastos de la vaina del haz donde es descarboxilado por la NADP-malato descarboxilasa (también conocida como enzima NADP málica) utilizando NADP como cofactor. La mayoría de las especies de gramíneas de Panicoideae utilizan el subtipo NADP-ME.
Subtipo NAD-ME, el CO2 se incorpora en el mesófilo y se transporta como aspartato a las células de la vaina del haz, donde se convierte en malato en las mitocondrias y se descarboxila mediante la NAD-malato descarboxilasa con NAD como cofactor. Este subtipo es característico de los miembros de Chloridoideae, incluidas las gramíneas de regiones cálidas, especialmente en hábitats secos.
Ecológicamente, como se describió anteriormente, existen diferencias en la distribución de los tres subtipos C4 . Sin embargo, el subtipo C4 no es necesariamente una adaptación a un clima particular y no es el único determinante de la distribución (D. J. Gibson, 2009).

Rutas metabólicas entre las células del mesofilo y las células de la vaina en los tres tipos de fotosíntesis C4 (D. J. Gibson, 2009).
Giussani et al. (2008) sugieren que la fotosíntesis C3 es la condición ancestral entre las gramíneas panicoides y que C4 surgió al menos ocho veces. Se desconoce el orden de evolución de los tres tipos.
El número de orígenes de la fotosíntesis C4 en Panicoideae es notable dada la aparente complejidad de la vía. La fotosíntesis de C4 requiere numerosas modificaciones bioquímicas y anatómicas de la planta, que aparentemente implican múltiples cambios genéticos, aunque, hasta donde se sabe, no hay genes o proteínas "nuevos" involucrados en las plantas C4. Las enzimas utilizadas son todas enzimas de mantenimiento C3 cuya regulación se altera de una manera específica para cada tejido.
Las distinciones entre la bioquímica de NAD-ME, PCK y NADP-ME no son tan marcadas como podría parecer. La enzima PCK sólo es activa en especies clasificadas como de tipo PCK, y casi no tiene actividad detectable en especies de tipo NAD-ME y NADP-ME , y la actividad de la enzima NADP-ME es muy baja en especies de tipo PCK . Sin embargo, el enzima descarboxilante de NAD-ME es activo no sólo en especies clasificadas como NAD-ME, sino también en especies de tipo PCK y en especies de tipo NADP-ME. En otras palabras, los subtipos C4 no reflejan en realidad distinciones absolutas en la actividad de la enzima descarboxilante. El malato es el principal ácido C4 formado en el mesófilo de las especies C4 NADP-ME, y el aspartato está presente principalmente en PCK y NAD-ME. Sin embargo, ambos productos se detectan en C3 y en todas las especies C4. Aunque la actividad de las enzimas difiere significativamente entre vías, células y orgánulos. Las vías evolutivas que conectan estos tres tipos deben haber pasado necesariamente por un "encendido" para la actividad descarboxilante de un tipo y un "apagado" simultáneo para la actividad de otro tipo.

No hay comentarios:
Publicar un comentario